以下文章来源于iNature
这些结构表明,在所有 Wnt 配体中都非常保守的 WNT3A 的发夹 2 从管腔侧插入 PORCN,而棕榈油酰辅酶 A 从胞质侧进入酶。催化组氨酸触发不饱和棕榈油酰基转移到 Wnt 发夹 2 上的目标丝氨酸,这得益于两个底物的接近。抑制剂结合结构表明 LGK974 占据了棕榈油酰辅酶 A 结合位点以阻止反应。因此,这项工作为 Wnt 酰化提供了一种机制,并推动了用于癌症治疗的 PORCN 抑制剂的开发。
 Wnt 信号是人类发育和维持干性的重要信号级联之一。Wnt 信号由分泌的脂质修饰信号糖蛋白 Wnt 与其受体 Frizzled 之间的相互作用触发。Porcupine (PORCN) 是内质网驻留膜结合 O-酰基转移酶 (MBOAT) 家族的成员,将棕榈油酰基部分转移到 Wnt 配体上的丝氨酸残基上。这种修饰对于 Wnt 信号激活是必不可少的,并且类似于 Hedgehog 信号转导中 Hedgehog 配体的棕榈酰修饰。结构研究表明,Wnt 和 Hedgehog 上的脂质修饰对于它们识别受体至关重要。
Wnt 信号是人类发育和维持干性的重要信号级联之一。Wnt 信号由分泌的脂质修饰信号糖蛋白 Wnt 与其受体 Frizzled 之间的相互作用触发。Porcupine (PORCN) 是内质网驻留膜结合 O-酰基转移酶 (MBOAT) 家族的成员,将棕榈油酰基部分转移到 Wnt 配体上的丝氨酸残基上。这种修饰对于 Wnt 信号激活是必不可少的,并且类似于 Hedgehog 信号转导中 Hedgehog 配体的棕榈酰修饰。结构研究表明,Wnt 和 Hedgehog 上的脂质修饰对于它们识别受体至关重要。 Wnt 上酰化位点的突变或 PORCN 活性的抑制消除了 Wnt 信号,表明 PORCN 介导的棕榈酰化在 Wnt 信号通路中的关键作用。迄今为止,已鉴定出通过旁分泌和自分泌信号通路激活 Wnt 信号通路的人类 Wnt 配体。相比之下,PORCN 是唯一催化 Wnt 脂化的酶,这暗示了 PORCN 介导的 Wnt 酰化的一般原理。
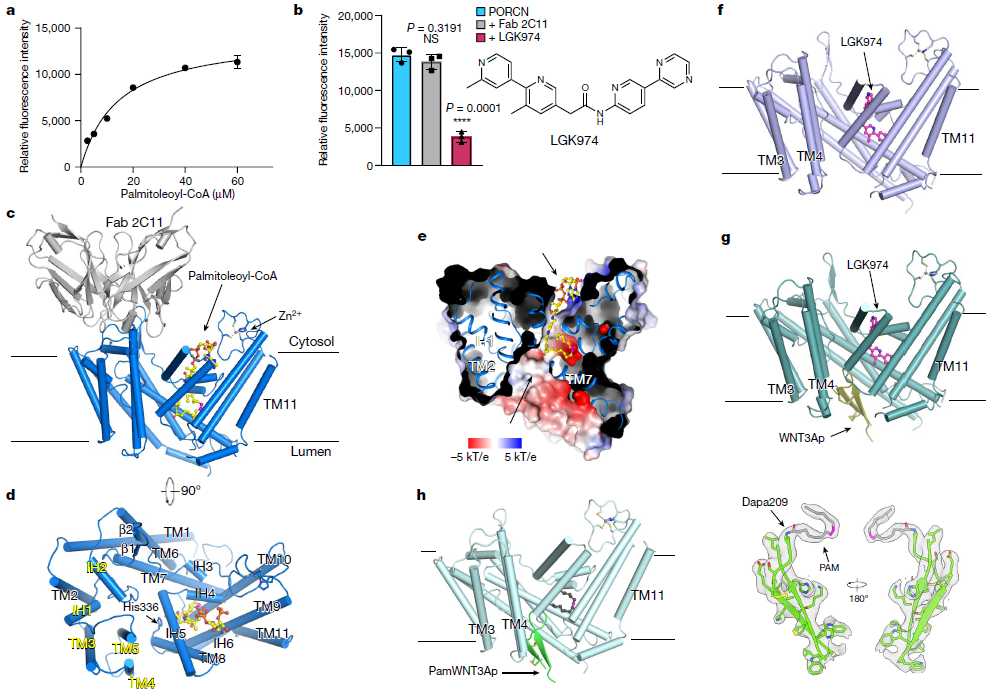
人类 PORCN 的功能表征和整体结构(图源自Nature )
MBOAT 将酰基从酰基辅酶或辅助蛋白质转移到脂质或蛋白质底物。该家族在脂质代谢和信号转导中具有重要作用。甾醇 O-酰基转移酶 (ACAT) 和甘油二酯 O-酰基转移酶 (DGAT) 的结构揭示了 MBOAT 介导的脂质修饰的机制。Hedgehog 酰基转移酶 (HHAT) 的结构研究表明,Hedgehog 的柔性 N 端肽插入 HHAT 中,从而在 Hedgehog 的 N 端半胱氨酸主链上添加了一个棕榈酰基。相比之下,Wnt 的脂质修饰发生在位于 Wnt 发夹 2 尖端的丝氨酸残基的羟基上。Wnt 发夹 2 的序列在不同 Wnt 之间高度保守,表明这种特殊的二级结构对于 PORCN 识别其底物是必不可少的。
PORCN 的缺乏及其突变导致局灶性真皮发育不全,这是一种与皮肤、手、脚和眼睛发育异常有关的罕见疾病。值得注意的是,已发现一些小分子抑制剂,包括 LGK974 和 ETC159,可特异性破坏 PORCN 酶活性,从而消除 Wnt 信号传导。这种抑制显著减小了 Wnt 依赖性肿瘤的大小。 LGK974目前正在进行治疗胰腺癌、黑色素瘤、三阴性乳腺癌和其他恶性肿瘤的临床试验。
在这里,该研究报告了 (1) PORCN 与棕榈酰辅酶 A 结合、(2) PORCN 与 LGK974 结合、(3) PORCN 与 WNT3A 发夹 2 和 LGK974 结合以及 (4) PORCN 的冷冻电子显微镜 (cryo-EM) 结构与合成的棕榈酰化 WNT3A 发夹 2 结合。这些结构阐明了 PORCN 介导的 Wnt 酰化的分子机制,并为开发小分子抗癌药物提供了见解。
李晓淳,现在美国西南医学中心分子遗传系任职。2004-2012年就读于清华大学获得理学学士和博士学位,导师施一公教授;2012-2017在美国洛克菲勒大学从事博士后研究,导师Günter Blobel为1999年诺贝尔生理学或医学奖得主。2017年6月就职于美国西南医学中心分子遗传系任助理教授,开展独立科研工作,利用结构生物学和生物化学手段研究胆固醇的生物合成和信号转导。在Science,Nature,PNAS 等(其中包含Science 2篇,Nature 8篇)发表文章达到40篇。
参考消息:
https://www.nature.com/articles/s41586-022-04952-2
https://www.utsouthwestern.edu/labs/li-xiaochun/
https://www.mbtechinst.qd.sdu.edu.cn/info/1074/1900.htm
题图:自制

好文不怕贵,舍得给稿费!
点击图片,可查看详细指南

我们还悄悄上线了「树洞」栏目,困扰,爆料,心事或者有趣的分享欢迎向我们砸来~
后台回复「树洞」,即可开始分享